




服(fu)務(wu)熱線(xian)
-
技(ji)術(shu)文章ARTICLE
您(nin)當前的位置(zhi):首頁(ye) > 技(ji)術(shu)文章 > FluorCam葉(ye)綠(lv)素熒光成(cheng)像(xiang)技(ji)術(shu):作(zuo)物(wu)基(ji)因功(gong)能與(yu)調(tiao)控(kong)機(ji)制(zhi)研(yan)究(jiu)FluorCam葉(ye)綠素熒光成(cheng)像(xiang)技(ji)術(shu):作(zuo)物(wu)基(ji)因功(gong)能與(yu)調(tiao)控(kong)機(ji)制(zhi)研(yan)究(jiu)
發(fa)布時間: 2021-09-08 點擊次數(shu): 3315次2021年中央(yang)壹(yi)號(hao)文件提出(chu)了解(jie)決(jue)種(zhong)源“卡脖(bo)子"問題的(de)要求。而開發(fa)更加(jia)優(you)質(zhi)高(gao)效的作(zuo)物(wu)品(pin)種(zhong)和(he)先(xian)進的(de)栽(zai)培(pei)技(ji)術(shu),解(jie)決(jue)種(zhong)源“卡脖(bo)子"問題,壹(yi)個(ge)非(fei)常(chang)重要的研究方(fang)向(xiang)是(shi)將植(zhi)物(wu)基(ji)因組(zu)學與(yu)表(biao)型組(zu)學進(jin)行(xing)結(jie)合來(lai)進(jin)行(xing)育種(zhong)工作(zuo)。壹(yi)方(fang)面,作(zuo)物(wu)基(ji)因的功(gong)能與(yu)調(tiao)控(kong)機(ji)制(zhi)必須(xu)進(jin)行(xing)相(xiang)關的(de)表(biao)型驗(yan)證,才能(neng)確(que)定其確(que)實(shi)具備(bei)相(xiang)應(ying)的(de)功(gong)能,比如光合能(neng)力變(bian)化(hua)、抗(kang)逆(ni)性調(tiao)控(kong)等(deng);另(ling)壹(yi)方(fang)面,也可(ke)以通(tong)過(guo)對(dui)優(you)良表(biao)型的(de)篩(shai)選(xuan),獲(huo)得(de)可(ke)穩定遺(yi)傳(chuan)的(de)基(ji)因型。
FluorCam葉(ye)綠(lv)素熒光成(cheng)像(xiang)技(ji)術(shu)作(zuo)為(wei)最重要的表(biao)型成(cheng)像(xiang)分析技(ji)術(shu)之(zhi)壹(yi),在(zai)作(zuo)物(wu)基(ji)因功(gong)能與(yu)調(tiao)控(kong)機(ji)制(zhi)研(yan)究(jiu)上都有(you)大(da)量應(ying)用。下面僅(jin)列舉2020年發(fa)表(biao)的部(bu)分文獻案(an)例(li):
1. 玉米(mi)固(gu)碳饑餓狀態下(xia)自體吞噬(shi)回收(shou)的多組(zu)學分析
聖路易(yi)斯(si)華盛頓大(da)學在(zai)研(yan)究玉米(mi)在(zai)固(gu)碳饑餓狀態下(xia)的自體吞噬(shi)回收(shou)過程(cheng)時應(ying)用(yong)了綜(zong)合多組(zu)學分析方(fang)法(fa),FluorCam葉(ye)綠素熒光成(cheng)像(xiang)技(ji)術(shu)作(zuo)為(wei)*代(dai)表(biao)性的光合表(biao)型研(yan)究技(ji)術(shu)正(zheng)適(shi)合這(zhe)壹(yi)研(yan)究。通(tong)過(guo)對(dui)核(he)心(xin)自體吞噬(shi)組(zu)分ATP12缺乏突變(bian)體(ti)的(de)多組(zu)學分析發(fa)現,固(gu)碳饑餓會造(zao)成氨(an)基(ji)酸、碳水化(hua)合物(wu)、核(he)酸相關代(dai)謝(xie)物(wu)的(de)極大(da)變(bian)化(hua),但通(tong)過(guo)FluorCam測量得(de)到(dao)最小葉(ye)綠(lv)素熒光F0和(he)最大(da)葉(ye)綠(lv)素熒光Fmax(Fm)則(ze)表(biao)明(ming),固(gu)碳饑餓只(zhi)會對(dui)光合作(zuo)用(yong)造(zao)成限度的影響(xiang)。這(zhe)壹(yi)研(yan)究成(cheng)果發(fa)表(biao)於(yu)2020年(nian)《the Plant Cell》。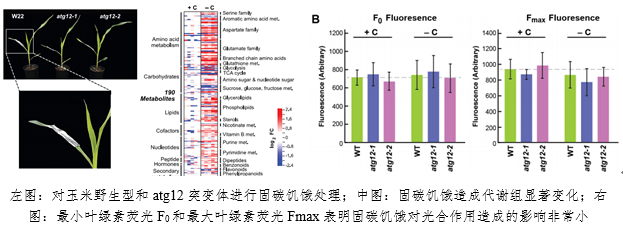
2. 水稻(dao)細(xi)胞(bao)分裂素受體(ti)對(dui)發(fa)育的調(tiao)節(jie)
細(xi)胞(bao)分裂素對植(zhi)物(wu)生(sheng)長(chang)發(fa)育的方(fang)方(fang)面面都有(you)調(tiao)節(jie)作(zuo)用(yong)。北(bei)卡羅來納大(da)學使(shi)用CRISPR-Cas9基(ji)因編輯(ji)技(ji)術(shu)破(po)壞(huai)了水(shui)稻(dao)的細(xi)胞(bao)分裂素組(zu)氨(an)酸激(ji)酶(mei)(HK)受體,從而研究(jiu)細(xi)胞(bao)分裂素在單(dan)子(zi)葉植(zhi)物(wu)中(zhong)的作(zuo)用(yong)。結(jie)果(guo)表(biao)明(ming),hk5和(he)hk6信(xin)號(hao)突變(bian)體(ti)影(ying)響(xiang)了根(gen)系(xi)生長(chang)、葉片(pian)寬(kuan)度、花(hua)序結(jie)構(gou)和(he)花(hua)的發(fa)育等(deng)。同(tong)時,運用FluorCam葉(ye)綠(lv)素熒光成(cheng)像(xiang)技(ji)術(shu)分析熒光衰減(jian)比率(lv)RFd,表(biao)明(ming)在經(jing)過3天(tian)暗處(chu)理(li)後,缺少外(wai)源細(xi)胞(bao)分裂素時,hk5和(he)hk6信(xin)號(hao)突變(bian)體(ti)的(de)RFd顯著(zhu)低(di)於(yu)野(ye)生(sheng)型;而在補(bu)充外(wai)源細(xi)胞(bao)分裂素時則(ze)表(biao)現出(chu)顯著(zhu)的(de)趨緩反應。從而證明(ming)細(xi)胞(bao)分裂素在水(shui)稻(dao)光合作(zuo)用(yong)中發(fa)揮重(zhong)要功(gong)能。這(zhe)壹(yi)研(yan)究發(fa)表(biao)於(yu)2020年(nian)《Development》。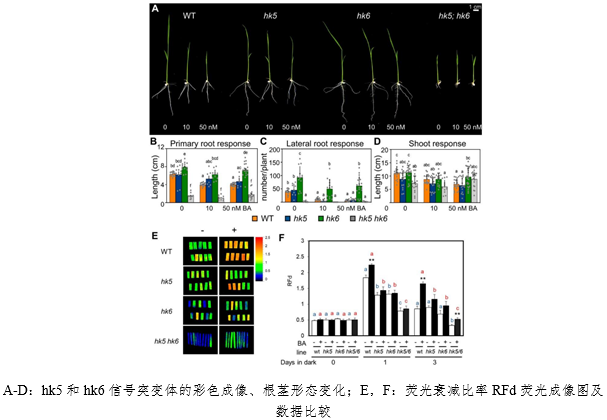
3. CBL4-CIPK5信號(hao)通(tong)路(lu)對(dui)水(shui)稻(dao)鹽抗(kang)性的調(tiao)節(jie)作(zuo)用(yong)
鈣調(tiao)神(shen)經(jing)磷(lin)酸酶B亞基(ji)樣(yang)蛋白(bai)(CBL)和(he)CBL互作(zuo)蛋白(bai)激(ji)酶(mei)(CIPK)在植(zhi)物(wu)非(fei)生物(wu)脅(xie)迫(po)調(tiao)控(kong)中(zhong)扮演重要角色。華南(nan)農業(ye)大(da)學通(tong)過(guo)轉(zhuan)基(ji)因方(fang)法(fa)使(shi)水稻(dao)過表(biao)達(da)CdtCIPK5和(he)CdtCBL4,然(ran)後(hou)對轉(zhuan)基(ji)因水稻(dao)進行(xing)鹽脅迫(po)、低(di)溫脅(xie)迫(po)和(he)幹(gan)旱脅迫(po)處(chu)理(li)。通(tong)過(guo)FluorCam葉(ye)綠(lv)素熒光成(cheng)像(xiang)系(xi)統測量的(de)最大(da)光化(hua)學效率Fv/Fm證(zheng)明(ming),CdtCIPK5和(he)CdtCBL4過(guo)表(biao)達(da)提高(gao)了水(shui)稻(dao)的鹽脅迫(po)抗(kang)性,但對低(di)溫和(he)幹(gan)旱脅迫(po)則沒(mei)有(you)顯著(zhu)作(zuo)用(yong)。這(zhe)壹(yi)研(yan)究發(fa)表(biao)於(yu)2020年(nian)《Environmental and Experimental Botany》。
4. PPR-SMR蛋白(bai)對(dui)水(shui)稻(dao)、玉米(mi)葉(ye)綠體mRNA的(de)編輯(ji)作(zuo)用(yong)
華中(zhong)農業(ye)大(da)學研(yan)究(jiu)了壹(yi)種(zhong)水(shui)稻(dao)突變(bian)體(ti)osatp4。這(zhe)種(zhong)突變(bian)體(ti)缺(que)乏(fa)壹(yi)種(zhong)玉米(mi)PPR-SMR蛋白(bai)ATP4的(de)同(tong)源基(ji)因。20℃低(di)溫處(chu)理(li)後,在表(biao)型上來說,osatp4突變(bian)體(ti)會(hui)表(biao)現出(chu)萎(wei)黃病。而通(tong)過(guo)FluorCam葉(ye)綠(lv)素熒光成(cheng)像(xiang)則(ze)證(zheng)明(ming),20℃低(di)溫下(xia),osatp4突變(bian)體(ti)的(de)最大(da)光化(hua)學效率Fv/Fm顯著(zhu)低(di)於(yu)野(ye)生(sheng)型。這(zhe)表(biao)明(ming)ATP4是與(yu)水稻(dao)的低(di)溫響(xiang)應(ying)和(he)光合機(ji)制(zhi)相(xiang)關的(de)。進(jin)壹(yi)步研究發(fa)現ATP4會(hui)促(cu)進(jin)葉綠體(ti)rps8 mRNA的編(bian)輯(ji)。這(zhe)壹(yi)研(yan)究成(cheng)果發(fa)表(biao)於(yu)2020年(nian)《Plant Physiology》。
參考(kao)文獻:
1.McLoughlin F, et al. 2020. Autophagy Plays Prominent Roles in Amino Acid, Nucleotide, and Carbohydrate Metabolism during Fixed-Carbon Starvation in Maize. The Plant Cell 32: 2699–2724
2.Burr CA, et al. 2020. The HK5 and HK6 cytokinin receptors mediate diverse developmental pathways in rice. Development 147: dev191734
3.Huang S, et al. 2020. CBL4-CIPK5 pathway confers salt but not drought and chilling tolerance by regulating ion homeostasis. Environmental and Experimental Botany 179: 104230
4.Zhang J, et al. 2020. The PPR-SMR protein ATP4 is required for editing the chloroplast rps8 mRNA in rice and maize. Plant Physiology 184: 2011–2021