




服務熱線(xian)
-
技術(shu)文(wen)章ARTICLE
您當(dang)前的(de)位(wei)置(zhi):首頁 > 技術(shu)文(wen)章 > 植(zhi)物(wu)表型(xing)成(cheng)像分析技術(shu)應(ying)用案(an)例(li)—雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)植(zhi)物(wu)表型(xing)成(cheng)像分析技術(shu)應(ying)用案(an)例(li)—雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)
發(fa)布(bu)時(shi)間(jian): 2025-08-07 點(dian)擊次(ci)數: 719次(ci)雜(za)種(zhong)優勢(heterosis,hybrid vigor)是(shi)雜(za)合(he)體(ti)在壹種(zhong)或多種(zhong)性狀上(shang)優於兩個(ge)親(qin)本(ben)的(de)現(xian)象(xiang)。例(li)如(ru)不(bu)同品(pin)系(xi)、不(bu)同品(pin)種(zhong)、甚至不(bu)同種(zhong)屬間進(jin)行雜(za)交所(suo)得(de)到(dao)的(de)雜(za)種(zhong)壹代(dai)往(wang)往(wang)比(bi)它(ta)的(de)雙(shuang)親(qin)表現(xian)更(geng)強(qiang)大(da)的(de)生長(chang)速率和(he)代(dai)謝(xie)功能,從(cong)而(er)導致(zhi)器(qi)官(guan)發(fa)達、體(ti)型(xing)增(zeng)大、產量(liang)提(ti)高,或者(zhe)表現(xian)在(zai)抗病、抗蟲、抗逆(ni)力(li)、成活(huo)力(li)、生殖(zhi)力(li)、生存力(li)等的(de)提(ti)高。利用雜(za)種(zhong)優勢進行雜(za)交育(yu)種(zhong)是(shi)作物(wu)育種(zhong)的(de)重(zhong)要手(shou)段之(zhi)壹。
 雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"/>
雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"/>雜(za)種(zhong)優勢作為作物(wu)育種(zhong)重(zhong)要表型(xing)性(xing)狀(zhuang)(traits),對其(qi)高通(tong)量分析檢測具有(you)特(te)別(bie)重(zhong)要的(de)現(xian)實(shi)意義(yi),下(xia)面我們(men)通過(guo)幾個(ge)研(yan)究(jiu)案(an)例(li),推薦出(chu)雜(za)交育(yu)種(zhong)表型(xing)分析檢測技術(shu)全(quan)面解決(jue)方(fang)案(an):
案例(li)壹、小麥(mai)—偃(yan)麥(mai)草雜(za)交系(xi)的(de)抗(kang)旱特(te)性
雜(za)種(zhong)優勢:幹(gan)旱脅迫下(xia)盡(jin)量(liang)保(bao)持(chi)葉(ye)綠(lv)素含(han)量(liang),減(jian)少葉(ye)片變(bian)黃,維(wei)持(chi)植(zhi)株(zhu)面積(ji)基(ji)本不(bu)變
小麥(mai)的(de)野生近緣種(zhong)Thinopyrum屬(特(te)別(bie)是(shi)中(zhong)間偃(yan)麥(mai)草Th. intermedium和長(chang)穗偃(yan)麥(mai)草Th. ponticum)具有(you)優異(yi)的(de)抗(kang)逆性。匈牙利農業(ye)研究(jiu)中(zhong)心的(de)科(ke)學(xue)家利用這(zhe)兩種(zhong)偃(yan)麥(mai)草的(de)雜(za)交種(zhong)Agropyron glel(GLAEL)來開(kai)發(fa)新(xin)型(xing)小麥(mai)抗(kang)性(xing)品(pin)種(zhong)。他們通(tong)過(guo)小麥(mai)品(pin)種(zhong)MV9與(yu)GLAEL雜(za)交,經多代(dai)回(hui)交(jiao)選(xuan)育出44條(tiao)染(ran)色(se)體(ti)的(de)附(fu)加(jia)系GLA7。
為了(le)評估GLA7的(de)抗(kang)旱優勢,研究(jiu)者(zhe)通過(guo)PlantScreen傳送帶(dai)式高通(tong)量表型(xing)成(cheng)像系統(tong),對GLA7和其(qi)親(qin)本(ben)小麥(mai)基(ji)因型(xing)MV9 和KAR進行了(le)幹(gan)旱培養和RGB形(xing)態表型(xing)成(cheng)像分析。PlantScreen系統利用(yong)自(zi)帶(dai)的(de)氣(qi)候控制(zhi)生長(chang)室(shi)在小麥(mai)的(de)分蘗(nie)期(qi)、抽(chou)穗期(qi)和(he)花(hua)期(qi)分別(bie)模(mo)擬了(le)不(bu)同的(de)晝(zhou)夜(ye)光照(包括(kuo)光強(qiang)、光質組成、光照周期(qi))、溫度(du)並(bing)自(zi)動(dong)進(jin)行稱重(zhong)澆灌。對(dui)照組壹直維(wei)持(chi)土壤(rang)含(han)水(shui)量(liang),處理(li)組則在(zai)花期(qi)開(kai)始前停止澆水10天。
 雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"style="width:800px;"/>
雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"style="width:800px;"/>結果表明,幹(gan)旱明顯(xian)造成(cheng)了(le)MV9 和KAR的(de)衰(shuai)老(lao),其(qi)葉(ye)片中(zhong)黃色(se)部分顯著(zhu)增(zeng)加。GLA7則明顯(xian)保(bao)持(chi)了(le)更(geng)多的(de)綠(lv)色(se)部分,說(shuo)明幹(gan)旱造成(cheng)的(de)其(qi)葉(ye)綠(lv)素降解要比它(ta)的(de)母(mu)本少得多。同(tong)時(shi),MV9 和KAR的(de)植(zhi)株(zhu)面積(ji)在(zai)幹(gan)旱脅迫處理(li)後(hou)顯(xian)著(zhu)降(jiang)低,而(er)GLA7的(de)植(zhi)株(zhu)面積(ji)則(ze)幾乎沒有(you)降低(di)。這(zhe)些(xie)表型(xing)數(shu)據(ju)充(chong)分證明了(le)GLA7在幹(gan)旱脅迫時(shi)的(de)雜(za)種(zhong)優勢。
案例(li)二(er)、通(tong)過(guo)葉(ye)綠(lv)素熒光成像技術(shu)研(yan)究(jiu)二(er)倍(bei)體(ti)馬(ma)鈴薯光合(he)特(te)性
雜(za)種(zhong)優勢:提高光合(he)效(xiao)率並(bing)增(zeng)加塊莖(jing)產量(liang)
馬(ma)鈴薯作為全球(qiu)重(zhong)要糧食(shi)作物(wu),其(qi)四(si)倍(bei)體(ti)遺傳復(fu)雜(za)性對(dui)育種(zhong)造成(cheng)了(le)壹定(ding)困難,而(er)二(er)倍(bei)體(ti)馬(ma)鈴薯為研究(jiu)提(ti)供(gong)了(le)簡化模(mo)型(xing)。荷蘭瓦格(ge)寧(ning)根(gen)大(da)學(xue)使用二(er)倍(bei)體(ti)馬(ma)鈴薯雜(za)交種(zhong)(如(ru)SOL015-3071和SOL014-7866)及其(qi)親(qin)本(ben)(D1、D16、DS),通過(guo)FluorCam葉(ye)綠(lv)素熒光成像技術(shu)分析二(er)倍(bei)體(ti)馬(ma)鈴薯在正(zheng)常培養條(tiao)件(jian)和(he)逆(ni)境(鹽脅迫、低溫、磷(lin)缺(que)乏(fa))下(xia)的(de)量(liang)子產額(e)ΦPSII(也(ye)稱為實(shi)際光化學(xue)效(xiao)率)遺(yi)傳變(bian)異(yi),用於評估其(qi)光合(he)效(xiao)率優勢,同時(shi)分析光合(he)效(xiao)率與(yu)塊莖(jing)產量(liang)的(de)相關性(xing)。
 雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"style="width:800px;"/>
雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"style="width:800px;"/>結果表明,在(zai)正(zheng)常生長(chang)狀(zhuang)態下(xia),雜(za)交種(zhong)的(de)ΦPSII顯著(zhu)高(gao)於其(qi)親(qin)本(ben),表現(xian)出(chu)明顯(xian)的(de)雜(za)種(zhong)優勢。同時(shi)幼苗期(qi)高(gao)ΦPSII(>0.6)的(de)植(zhi)株(zhu)在(zai)成熟期(qi)塊莖(jing)幹(gan)重(zhong)顯著(zhu)更(geng)高(24.3 g vs. 15 g)。
 雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"/>
雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"/>而在脅迫條(tiao)件(jian)下(xia),低(di)溫和(he)磷缺(que)乏(fa)顯著(zhu)降(jiang)低了(le)兩(liang)種(zhong)雜(za)交種(zhong)的(de)ΦPSII,其(qi)中(zhong)低溫的(de)降(jiang)幅更大。鹽脅迫未顯著(zhu)影響ΦPSII,但植(zhi)株(zhu)生長(chang)受(shou)限,FluorCam葉(ye)綠(lv)素熒光成像技術(shu)可高效(xiao)篩(shai)選(xuan)馬(ma)鈴薯的(de)早期(qi)逆(ni)境響應(ying)。
案例(li)三(san):擬南(nan)芥(jie)生物(wu)量雜(za)種(zhong)優勢的(de)生理機制(zhi)
雜(za)種(zhong)優勢:提高萌(meng)發(fa)率和(he)葉(ye)片早期(qi)發(fa)育(yu),維(wei)持(chi)光合(he)效(xiao)率不(bu)變,從(cong)而(er)提(ti)高生物(wu)量
擬南(nan)芥(jie)Arabidopsis thaliana雜(za)交種(zhong)往(wang)往(wang)會(hui)表現(xian)出(chu)生物(wu)量增(zeng)加的(de)雜(za)種(zhong)優勢。澳大利亞(ya)聯(lian)邦科學(xue)和(he)工業研(yan)究(jiu)組織與(yu)悉尼(ni)科(ke)技(ji)大(da)學(xue)合(he)作,旨(zhi)在確定(ding)擬南(nan)芥(jie)雜(za)交種(zhong)生物(wu)量增(zeng)加是(shi)由於單(dan)位(wei)葉(ye)面積(ji)光合(he)效(xiao)率提(ti)高,還是(shi)總體(ti)葉(ye)面積(ji)增(zeng)加導(dao)致總光合(he)產物(wu)增(zeng)多。
 雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"/>
雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"/>研究(jiu)人(ren)員通(tong)過(guo)FluorCam葉(ye)綠(lv)素熒光成像系統(tong)的(de)light curve光響應(ying)曲(qu)線測量程(cheng)序,測量了(le)雜(za)交種(zhong)和親(qin)本(ben)在(zai)不(bu)同光強(qiang)下(xia)的(de)量(liang)子產額(e)ΦPSII與(yu)電子傳遞速率ETR,並(bing)進(jin)壹步擬合(he)出(chu)最(zui)大(da)電(dian)子傳遞速率Jmax(即ETRmax)。結果表明雜(za)交種(zhong)和親(qin)本(ben)在(zai)光系統(tong)II效(xiao)率方(fang)面沒有(you)顯著(zhu)差(cha)異(yi)。
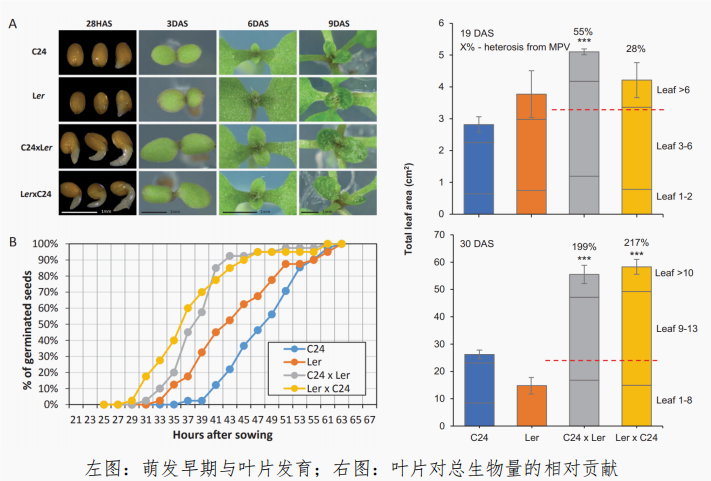 雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"/>
雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"/>而萌(meng)發(fa)與(yu)培養實(shi)驗表明,雜(za)交種(zhong)比親(qin)本(ben)更早萌(meng)發(fa),而(er)且在早期(qi)發(fa)育(yu)階段葉(ye)片比(bi)親(qin)本(ben)要大,進(jin)而(er)在(zai)生長(chang)後(hou)期(qi)獲(huo)取了(le)更(geng)高的(de)總葉(ye)面積(ji)。研(yan)究(jiu)得(de)到(dao)的(de)最(zui)終(zhong)結論(lun)是(shi)擬南(nan)芥(jie)雜(za)交種(zhong)的(de)生物(wu)量雜(za)種(zhong)優勢主要源於更大的(de)葉(ye)面積(ji)而非更高(gao)的(de)光合(he)效(xiao)率,萌(meng)發(fa)和(he)葉(ye)片早期(qi)發(fa)育(yu)則是(shi)關鍵(jian)驅動(dong)因(yin)素。光合(he)作用單(dan)位(wei)葉(ye)面積(ji)效(xiao)率不(bu)變,但(dan)總光合(he)產物(wu)增(zeng)加推動(dong)了(le)雜(za)交種(zhong)的(de)生長(chang)優勢。
案例(li)四(si):幹(gan)旱條(tiao)件(jian)下(xia)玉(yu)米雜(za)交種(zhong)的(de)不(bu)利因(yin)素
雜(za)種(zhong)優勢:在非脅迫條(tiao)件(jian)下(xia)有(you)較大(da)的(de)植(zhi)物(wu)體(ti)型(xing),但(dan)幹(gan)旱脅迫下(xia)光合(he)系(xi)統(tong)受(shou)損也(ye)更為嚴重(zhong)
這(zhe)壹研究(jiu)案(an)例(li)探(tan)討了(le)兩(liang)種(zhong)玉米自(zi)交(jiao)系(xi)及(ji)其(qi)F1雜(za)交種(zhong)在幹(gan)旱條(tiao)件(jian)下(xia)的(de)表現(xian)差(cha)異(yi)。研究(jiu)團隊來自(zi)捷克(ke)的(de)多所(suo)大(da)學(xue)和(he)研究(jiu)機構(gou),包括(kuo)查(zha)爾斯(si)大(da)學(xue)和(he)捷克(ke)科學(xue)院(yuan)實(shi)驗植(zhi)物(wu)學(xue)研(yan)究(jiu)所(suo)。
研究(jiu)中(zhong)對玉(yu)米光合(he)作用的(de)測量使用了(le)LCpro便(bian)攜式光合(he)儀(yi)和(he)FluorPen手(shou)持(chi)式葉(ye)綠(lv)素熒光儀。光合(he)儀(yi)測量數(shu)據(ju)包括凈(jing)光合(he)速率(Pn)、蒸騰(teng)速率(E)、氣(qi)孔導度(gs)、水分利用效(xiao)率(WUE)等。葉(ye)綠(lv)素熒光儀測量OJIP快速葉(ye)綠(lv)素熒光動(dong)力(li)學(xue)曲(qu)線,並(bing)計算(suan)了(le)最(zui)大量子產額(e)φP0、能量耗散量(liang)子產額(e)φD0、性能指數(shu)PIABS和PITOTAL等。
 雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"style="width:800px;"/>
雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"style="width:800px;"/>研究(jiu)結果表明,玉(yu)米雜(za)交種(zhong)在非脅迫條(tiao)件(jian)下(xia)表現(xian)出(chu)的(de)正(zheng)雜(za)種(zhong)優勢(較大(da)的(de)植(zhi)物(wu)體(ti)型(xing))在(zai)水(shui)分受限時(shi)成(cheng)為明顯(xian)劣(lie)勢,包括光系統(tong)各(ge)方(fang)面的(de)性(xing)能與(yu)效(xiao)率比(bi)親(qin)本(ben)有(you)更顯(xian)著(zhu)的(de)降(jiang)低、氣(qi)孔導度進壹步降(jiang)低(di)從而(er)造成(cheng)的(de)蒸(zheng)騰(teng)與(yu)凈(jing)光合(he)速率減(jian)低等。這(zhe)壹研究(jiu)強(qiang)調(tiao)了(le)植(zhi)物(wu)體(ti)型(xing)大(da)小與(yu)抗旱性的(de)關(guan)系,指出了(le)光合(he)系(xi)統(tong)不(bu)同組分功能在(zai)幹(gan)旱響應(ying)中(zhong)的(de)關(guan)鍵作用,為未來玉(yu)米抗旱育種(zhong)策(ce)略(lve)的(de)制(zhi)定(ding)提供了(le)理(li)論(lun)基(ji)礎和(he)實(shi)踐指(zhi)導(dao)。
LCpro光合(he)儀(yi)和(he)FluorPen手(shou)持(chi)式葉(ye)綠(lv)素熒光儀都(dou)具備小巧輕(qing)便(bian)、功能全(quan)面、堅固耐用(yong)等(deng)優點(dian)。FluorPen還可同時(shi)進(jin)行PAM脈沖(chong)調試式葉(ye)綠(lv)素熒光和OJIP快速葉(ye)綠(lv)素熒光的(de)測量,這(zhe)在目前所(suo)有(you)野外(wai)便(bian)攜式葉(ye)綠(lv)素熒光儀中(zhong)都可說(shuo)是(shi)獨樹壹幟的(de)。這(zhe)兩款(kuan)儀(yi)器(qi)在(zai)國內(nei)外(wai)都有(you)大量(liang)的(de)科(ke)研案例(li),這(zhe)兩款(kuan)儀(yi)器(qi)的(de)組合(he)更(geng)可以完(wan)整(zheng)測量植(zhi)物(wu)光合(he)作用過(guo)程與(yu)機制(zhi),是(shi)植(zhi)物(wu)光合(he)效(xiao)率測量、逆(ni)境脅迫評估中(zhong)的(de)科(ke)研利器(qi)。我們(men)有(you)幾百篇科相關研(yan)論(lun)文利(li)用可供有(you)關科(ke)研工作者(zhe)參考。
 雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"style="width:800px;"/>
雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"style="width:800px;"/>北京易科(ke)泰(tai)生態技術(shu)公(gong)司提供雜(za)交育(yu)種(zhong)表型(xing)分析技術(shu)全(quan)面解決(jue)方(fang)案(an)並(bing)提(ti)供(gong)相關參(can)考文獻:
Ø 國際PlantScreen植(zhi)物(wu)高通(tong)量(liang)表型(xing)成(cheng)像分析系統(tong),有(you)傳送帶(dai)版、XYZ版、PlantScreen SC、根系表型(xing)分析等不(bu)同功能規(gui)格供(gong)選(xuan)配(pei)
Ø PhenoTron®植(zhi)物(wu)表型(xing)成(cheng)像分析系統(tong),易(yi)科(ke)泰(tai)新(xin)壹代(dai)農業(ye)傳感(gan)器(qi)技(ji)術(shu)+AI平臺技術(shu),可根據國(guo)內(nei)用戶(hu)實(shi)際需(xu)求(qiu)及安裝場(chang)景(jing)靈活配(pei)置(zhi)定(ding)制(zhi),具備高(gao)光譜成(cheng)像、UV-MCF紫(zi)外(wai)光激發(fa)植(zhi)物(wu)熒光(脅迫誘(you)導次(ci)級代(dai)謝(xie)產物(wu)熒光)高光譜成(cheng)像、葉(ye)綠(lv)素熒光成像、Thermo-RGB紅外(wai)熱成(cheng)像與(yu)RGB成像融合(he)分析、3D激光掃(sao)描(miao)成像分析等表型(xing)分析技術(shu),AI平臺技術(shu)包(bao)括自(zi)動(dong)傳送系統(tong)、XYZ平臺、懸(xuan)浮(fu)雙(shuang)規(gui)平臺(tai)、機器(qi)人(ren)平臺等(deng)不(bu)同智(zhi)能(neng)平(ping)臺供選(xuan)配(pei)
 雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"style="width:800px;"/>
雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"style="width:800px;"/>Ø FluorCam葉(ye)綠(lv)素熒光成像系統(tong),有(you)幾千(qian)篇國(guo)際(ji)科(ke)研文獻可供參考
Ø FluorTron®植(zhi)物(wu)光合(he)表型(xing)成(cheng)像分析系統(tong),可根據客戶(hu)需(xu)求(qiu)定(ding)制(zhi)不(bu)同成(cheng)像面積(ji)的(de)葉(ye)綠(lv)素熒光動(dong)態(tai)成(cheng)像、葉(ye)綠(lv)素熒光光譜成(cheng)像、LEDIF(植(zhi)物(wu)群體(ti))冠層(ceng)葉(ye)綠(lv)素熒光成像系統(tong)等
 雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"style="width:800px;"/>
雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"style="width:800px;"/>Ø PhenoTron®智能LED光源培(pei)養(yang)與(yu)表型(xing)分析平臺(tai),集多通(tong)道(dao)智(zhi)能LED光源與(yu)多源表型(xing)傳感(gan)器(qi)技(ji)術(shu)於壹身(shen),可在對植(zhi)物(wu)進行精確光照培訓(xun),同時(shi)實(shi)現葉(ye)綠(lv)素(含(han)量(liang))與(yu)葉(ye)綠(lv)素熒光成像、Thermo-RGB成像、多功能高(gao)光譜成(cheng)像等在(zai)線晝(zhou)夜(ye)全天候監測分析植(zhi)物(wu)表型(xing)。
 雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"/>
雜(za)種(zhong)優勢與(yu)雜(za)交育(yu)種(zhong)"/>Ø PhenoTron®智能LED光源培(pei)養(yang)與(yu)表型(xing)分析平臺(tai)與(yu)西洋參誘(you)變育種(zhong)葉(ye)綠(lv)素熒光成像(EcoTech®實(shi)驗室)
Ø FluorPen系列(lie)手(shou)持(chi)式葉(ye)綠(lv)素熒光儀(有(you)探(tan)頭(tou)式和葉(ye)夾式,藻(zao)類(lei)用(yong)AquaPen試管(guan)式葉(ye)綠(lv)素熒光儀),可與(yu)LCi、LCpro等光合(he)儀(yi)組成便(bian)攜式大田作物(wu)表型(xing)分析測量系(xi)統(tong)
Ø FluorTron系列(lie)多光譜葉(ye)綠(lv)素熒光成像/多功能高(gao)光譜成(cheng)像分析系統(tong)
Ø PhenoPlot®輕(qing)便(bian)型(xing)大(da)田或溫室(shi)作物(wu)表型(xing)成(cheng)像分析系統(tong)
Ø PhenoTron® PTS植(zhi)物(wu)表型(xing)成(cheng)像分析系統(tong)
Ø 田間(jian)智(zhi)能巡檢(jian)機器(qi)人(ren)表型(xing)分析平臺(tai)
Ø ET-LEDIF葉(ye)綠(lv)素熒光監測系統(tong)
Ø 模(mo)塊式作物(wu)表型(xing)成(cheng)像分析系統(tong)
參考文獻:
1. Kruppa K, Türkösi E, Holušová K, et al. Genotyping-by-sequencing uncovers a Thinopyrum 4StS· 1JvsS Robertsonian translocation linked to multiple stress tolerances in bread wheat. Theoretical and Applied Genetics, 2025, 138(1): 13.
2. Prinzenberg A E, Víquez‐Zamora M, Harbinson J, et al. Chlorophyll fluorescence imaging reveals genetic variation and loci for a photosynthetic trait in diploid potato[J]. Physiologia Plantarum, 2018, 164(2): 163-175.
3. Liu P C, Peacock W J, Wang L, et al. Leaf growth in early development is key to biomass heterosis in Arabidopsis[J]. Journal of experimental botany, 2020, 71(8): 2439-2450.
4. Holá D, Benešová M, Fischer L, et al. The disadvantages of being a hybrid during drought: A combined analysis of plant morphology, physiology and leaf proteome in maize[J]. PLoS One, 2017, 12(4): e0176121.